
Whatman 滤膜 10373870 34 GLASS 2.5IN 100/PK 34 GLASS 2.5IN 100/PK
日度归档:2024年3月11日
Pall颇尔滤膜 4672 FILTER ACRO 50, 6MHDC II 18/PK
Pall颇尔滤膜 4672 FILTER ACRO 50, 6MHDC II 18/PK 454 3634
康宁corning 4810 TIPS,ISOTIP,FILTER,1-200UL,S,R 96/包
康宁corning 4810 TIPS,ISOTIP,FILTER,1-200UL,S,R 96/包 10包/箱 2222.47
OXOID培养基 R455302 结晶紫中性红胆盐葡萄糖琼脂 Violet Red Bile Glucose Agar
OXOID培养基 R455302 结晶紫中性红胆盐葡萄糖琼脂 Violet Red Bile Glucose Agar 781.3468
OXOID培养基 SR0090A VITO+水华液 VITOX SUPP+RECONSTITUTION FLUID 2X5VIA
OXOID培养基 SR0090A VITO+水华液 VITOX SUPP+RECONSTITUTION FLUID 2X5VIA 261.63574976
1,3-?-D-葡聚糖内切酶检测底物 AZCL-Curdlan 3g 货号:I-AZCUR Megazyme中文站
1,3-?-D-葡聚糖内切酶检测底物
英文名:AZCL-Curdlan 3g
货号:I-AZCUR
规格:3g
1,3-?-D-葡聚糖内切酶检测底物
PDF Download
暂无问题解答
暂无视频
a-L-阿拉伯呋喃糖酶[杆菌芽胞] α-L-Arabinofuranosidase (Clostridium thermocellum) 货号:E-ABFCT Megazyme中文站
a-L-阿拉伯呋喃糖酶[杆菌芽胞]
英文名:α-L-Arabinofuranosidase (Clostridium thermocellum)
货号:E-ABFCT
规格:500 Units at 40°C
High purity recombinant alpha-L-Arabinofuranosidase (Clostridium thermocellum) for use in research, biochemical enzyme assays and in vitro diagnostic analysis.
EC 3.2.1.55
CAZy family: GH51
Recombinant. From Clostridium thermocellum.
In 3.2 M ammonium sulphate.
Specific activity: 58 U/mg protein on p-nitrophenyl-α-L-arabinofuranoside at pH 5.5 and 40oC (155 U/mg at pH 5.5 and 60oC).
Stable at 4oC for ~ 4 years.
暂无问题解答
暂无视频
康宁corning 4975 PIPETTE,1ML,ASPIRATION,S,BK,50 真空移液管 1ml 灭菌 独立包装 50个/包
康宁corning 4975 PIPETTE,1ML,ASPIRATION,S,BK,50
真空移液管 1ml 灭菌 独立包装 50个/包 20包/箱 2956.77
BIOLIKE® 构象分析ELISA试剂盒
| 产品编号 | 产品名称 | 产品规格 | 产品等级 | 产品价格 |
| ENZ-KIT118-0003 | BIOLIKE® Bevacizumab conformational ELISA kit 生物仿制药 贝伐单抗构建ELISA试剂盒 |
3×96 wells | - | - |
| ENZ-KIT119-0003 | BIOLIKE® Alemtuzumab conformational ELISA kit 生物仿制药 阿仑单抗构象ELISA试剂盒 |
3×96 wells | - | - |
| ENZ-KIT121-0003 | BIOLIKE® Trastuzumab conformational ELISA kit 生物仿制药 曲妥珠单抗构象ELISA试剂盒 |
3×96 wells | - | - |
| ENZ-KIT122-0003 | BIOLIKE® Adalimumab conformational ELISA kit 生物仿制药 阿达木单抗构建ELISA试剂盒 |
3×96 wells | - | - |
| ENZ-KIT123-0003 | BIOLIKE® Conserved Ab sequence conformational ELISA kit 生物仿制药 保守单抗顺序构象ELISA试剂盒 |
3×96 wells | - | - |
| ENZ-KIT124-0003 | BIOLIKE® Infliximab conformational ELISA 生物仿制药 英夫利昔单抗构建ELISA试剂盒 |
3×96 wells | - | - |
| ENZ-KIT125-0003 | BIOLIKE® Rituximab conformational ELISA kit 生物仿制药 利妥昔单抗构建ELISA试剂盒 |
3×96 wells | - | - |
| ENZ-KIT126-0003 | BIOLIKE® Palivizumab conformational ELISA kit 生物仿制药 帕利珠单抗构象ELISA试剂盒 |
3×96 wells | - | - |
|
构象分析ELISA试剂盒
|
|
此试剂盒可用于生物仿制药和投入市场的单克隆抗体之间的3-D构象比较和高级结构鉴定,这项新技术提供了一种在分子水平上对生物仿制药构象可比性的高灵敏度、系统性和稳定检测。 |

|
◆适用领域 ● 单克隆抗体 ● 药物偶联物 ● 细胞系筛选 ● 工艺过程开发 ● 剂型开发 ● 可比性研究 ● 产品特性分析 |
|

|
◆作用原理 该试剂盒采用多于三十个多克隆抗体覆盖一个单克隆抗体,来检测表面抗原表位的分布,系统性更强,灵敏度更高,提供用于生物仿制药和单克隆抗体高级结构可比性研究的独特方法。 采用抗生物体内全长蛋白序列的抗体,每个抗体都能特异性识别相应的多肽抗原。当多肽是正确折叠的全长蛋白时,其抗原性表现在3D结构中,只有有限数量的抗体会应答。 抗体结合部位可作为判定生物仿制药多肽正确折叠的指纹指标,如果生物仿制药正确折叠和糖基化,指纹便会符合。如果生物仿制药没有正确折叠,高级结构发生不同程度的改变时,原本被掩盖的抗原表位暴露出来,导致抗体对其识别的信号增强。 |
◆主要特点
● 生物仿制药和单克隆抗体之间的高级结构可比性的独特检测。
● 可检测到少量的构象杂质,含量低至0.1%。
● 支持后续的生物测定和糖基化分析。

BD培养基 217110 25 g EA Difco D-Mannose D-甘露糖
BD培养基 217110 25 g EA Difco D-Mannose D-甘露糖 
Whatman 滤膜 10539530 903 NACO HIV CARD 100/PK
Whatman 滤膜 10539530 903 NACO HIV CARD 100/PK 903 NACO HIV CARD 100/PK
康宁corning 430516 FLTR SYS,1000ML,.45UM,CA,S,IND 真空过滤系统 1000ml 0.45umCA(醋酸纤维)膜 90mm直径 灭菌 单个包装 1个/包
康宁corning 430516 FLTR SYS,1000ML,.45UM,CA,S,IND
真空过滤系统 1000ml 0.45umCA(醋酸纤维)膜 90mm直径 灭菌 单个包装 1个/包 12包/箱 3177.21
Beta葡聚糖[混联]检测试剂盒 β-Glucan Assay Kit (Mixed Linkage) 货号:K-BGLU Megazyme中文站
Beta葡聚糖[混联]检测试剂盒
英文名:β-Glucan Assay Kit (Mixed Linkage)
货号:K-BGLU
规格:100 assays per kit
分析物意义: 大麦和燕麦的主要细胞壁多糖
Megazyme检测试剂盒优点:反应迅速、试剂稳定, 只有酶检测试剂盒可用。 AOAC方法 995.16; AACC 方法 32-23
AACC Method 32-23.01, AOAC Method 995.16, EBC Methods 3.11.1, 4.16.1 and 8.11.1, ICC Standard No. 166 and RACI standard method for the measurement of 1,3:1,4-ß-D-glucan in cereal grains, milling fractions, wort and beer. Recommended/Standard procedure of the Association of Official Analytical Chemists (AOAC), American Association of Cereal Chemists (AACC), Royal Australian Chemical Institute (RACI), European Brewing Convention (EBC), and International Association for Cereal Science and Technology (ICC). Content:100 assays per kit
Colourimetric method for the determination of β-Glucan in
cereal grains, feed, foodstuffs, beverages and other materials
Principle:
(lichenase)
(1) β-Glucan + H2O → β-gluco-oligosaccharides
(β-glucosidase)
(2) β-Gluco-oligosaccharides + H2O → D-glucose
(glucose oxidase)
(3) D-Glucose + H2O + O2 → D-gluconate + H2O2
(peroxidase)
(4) 2H2O2 + p-hydroxybenzoic acid + 4-aminoantipyrine →
quinoneimine + 4H2O
Kit size: 100 assays
Method: Spectrophotometric at 510 nm
Total assay time: ~ 100 min
Detection limit: 0.5-100% of sample weight
Application examples:
Oats, barley, malt, wort, beer, food and other materials
Method recognition:
AOAC (Method 995.16), AACC (Method 32-23.01), EBC (Methods
3.10.1, 4.16.1 and 8.13.1), ICC (Standard No. 166), RACI (Standard
Method) and CODEX (Type II Method)
Advantages
- Very cost effective
- All reagents stable for > 2 years as supplied
- Only enzymatic kit available
- Very specific
- Simple format
- Mega-Calc™ software tool is available from our website for hassle-free raw data processing
- Standard included
Q1. Should the pH of the sample be adjusted even for samples in acidic media?
The pH of the assay solution after the sample is added should be the same as that of the assay buffer that is supplied with the kit.
Low sample volumes (e.g. 0.1 mL) are not likely to affect the pH of the assay solution and therefore may not require pH adjustment.
Samples above 0.1 mL are more likely to affect the pH of the assay solution and therefore the pH of these samples should be adjusted as described in the data booklet, prior to addition to the assay.
Q2. There is an issue with the performance of the kit; the results are not as expected.
If you suspect that the Megazyme test kit is not performing as expected such that expected results are not obtained please do the following:
- Ensure that you have tested the standard sample that is supplied with the Megazyme test kit.
- Send the results of the kit standard, blank samples and the results obtained for your sample, in the relevant MegaCalc spreadsheet (if available) to Megazyme (cs@megazyme.com). Where available the relevant MegaCalc spreadsheet can be downloaded from where the product appears on the Megazyme website.
- State the kit lot number being used (this is found on the outside of the kit box).
- State which assay format was used (refer to the relevant page in the kit booklet if necessary).
- State exact details of any modifications to the standard procedure that is provided by Megazyme.
- State the sample type and describe the sample preparation steps if applicable.
Q3. We have measured the molecular weight of beta-glucan originating from barley ordered from your company. Can you please tell us which method have you used for measuring molecular weight?
The MW’s were determined by Multiangle laser light scattering technique.
Q4. I have purchased barley beta-glucan (lot 30108) and carob galactomannan low viscosity (lot 30702) and would like to know what else there might be in these substrates.
Barley beta-glucan lot 30108 would contain about 3-4% arabinoxylan (this was produced in 1993). Material supplied post 1995 contains < 0.5% arabinoxylan. Carob galactomannan is quite pure (> 96%).
Q5. Although not specified on the beta-glucan data sheet, is the ratio of 1-3 to 1-4 bonds measured? If so, what is the ratio and would you expect it to remain standardised over a number of different batches?
The content of 1,3 bonds in barley beta-glucan is about 32% (from literature). We would not expect this to change much (if at all) over different batches.
Q6. Both substrates; barley beta-glucan and wheat arabinoxylan, are standardised to a specific viscosity, e.g. 23-24 cSt. Are the substrates adjusted to give this viscosity?
The substrates are enzymically treated to yield the required viscosity (20 ~ 30 cSt). Modification of the viscosity of beta-glucan is required for IOB viscometric malt beta-glucanase test (which works well).
Q7. We are setting up reducing-sugar assays for both beta-glucanase and xylanase from different fungal origins. Which barley beta-glucan do you recommend for the glucanase assay.
For glucanase assay we recommend the medium viscosity beta-glucan. We now offer low, medium and high viscosity wheat arabinoxylans, and we think that the low viscosity material will be best (easiest) to use in the reducing-sugar assay.
荧光染料63520 Pyrenebutyric acid NHS ester 100 mg
lumiprobe荧光染料63520 Pyrenebutyric acid NHS ester 100 mg lumiprobe荧光染料Pyrenebutyric acid 羧基活性酯 100 mg 110
荧光染料13580 Pyrene maleimide 1 mg
lumiprobe荧光染料13580 Pyrene maleimide 1 mg lumiprobe荧光染料Pyrene 马来酰亚胺基 1 mg 110
NISSUI 日本日水培养基 05816 尼克酸定量用基础培养基 Niacin Assay Medium 50g
NISSUI 日本日水培养基 05816 尼克酸定量用基础培养基 Niacin Assay Medium 50g

康宁corning 3387 TRANSWELL,96W, HTS,W/5.0uM MEM HTS Transwell-96膜嵌套 5.0um孔径PC(聚碳酸酯)膜带盖接收板 大包装 4个/包
康宁corning 3387 TRANSWELL,96W, HTS,W/5.0uM MEM
HTS Transwell-96膜嵌套 5.0um孔径PC(聚碳酸酯)膜带盖接收板 大包装 4个/包 2包/箱 10250.02
黄递酶[大肠杆菌] Diaphorase (E. coli) 货号:E-DIAEC Megazyme中文站
黄递酶[大肠杆菌]
英文名:Diaphorase (E. coli)
货号:E-DIAEC
规格:1000 Units
High purity Diaphorase (E. coli) for use in research, biochemical enzyme assays and in vitro diagnostic analysis.
EC 1.8.1.4
From E. coli.
In 3.2 M ammonium sulphate.
Specific activity: 69 U/mg (25oC, pH 9.0).
Stable at 4oC for > 2 years.
暂无问题解答
暂无视频
Amresco 0523-100G LITHIUM HYDROXIDE 氢氧化锂
Amresco 0523-100G LITHIUM HYDROXIDE 氢氧化锂
OXOID培养基 CM0077B 肝肉汤Liver Broth LIVER BROTH 500G
OXOID培养基 CM0077B 肝肉汤Liver Broth LIVER BROTH 500G 938.4340601664
Whatman 滤膜 111112 Nuclepore径迹蚀刻膜 圆型, 47mm 3.0m 100/盒
Whatman 滤膜 111112 Nuclepore径迹蚀刻膜 圆型, 47mm 3.0m 100/盒 NUC PC 47MM 3.0uM 100/PK
Sigma特殊产品Thiopropyl–Sepharose® 6B
Sigma特殊产品Thiopropyl–Sepharose® 6B
产品编号 T8387 | CAS号 68517-67-9 | SIGMA
包装:1, 5, 15 g in poly bottle
Physical form
Lyophilized powder stabilized with lactose and dextran
Application
Thiopropyl Sepharose® 6B is used in protein chromatography, affinity chromatography and hydrophobic interactions. Thiopropyl Sepharose® 6B has been used for the removal of inhibitors from PCR extracts from evidence collected for law enforcement murder investigations. Thiopropyl Sepharose® 6B has also been used to describe a method for the investigation of functional properties of distinct domains of viral thiol proteins, including the influenza virus membrane M1 protein.
法律信息
Sepharose is a registered trademark of GE Healthcare
Pall颇尔滤膜 66609 FILTER DISC 0.45UM 90MM PK100
Pall颇尔滤膜 66609 FILTER DISC 0.45UM 90MM PK100 998 7985
Whatman 滤膜 10549684 FT020 150x150MM 10/PK
Whatman 滤膜 10549684 FT020 150x150MM 10/PK FT020 150x150MM 10/PK
Amresco 0305-2.5KG AMMONIUM CARBONATE 碳酸铵
Amresco 0305-2.5KG AMMONIUM CARBONATE 碳酸铵
有机锡化合物分析用|环境分析纯试剂(其他)|环境分析纯试剂|试剂|关东化学株式会社
小册子
该文章由WP-AutoPost插件自动采集发布
康宁corning 431153 FLTR SYS,150ML,.22UM,PES,S,IND 真空过滤系统 150ml 0.22um PES(聚醚砜)膜 50mm直径 灭菌 单个包装9(cf 431153/431154/431155/430516) 1个/包
康宁corning 431153 FLTR SYS,150ML,.22UM,PES,S,IND
真空过滤系统 150ml 0.22um PES(聚醚砜)膜 50mm直径 灭菌 单个包装9(cf 431153/431154/431155/430516) 1个/包 12包/箱 1226.87
Ready PCR Mix AMRESCO N806
名称:Ready PCR Mix
品牌:AMRESCO
订货号:N806

咨询此产品
产品介绍
Ready PCR Mix is a PCR master mix that enables reaction set-up through product visualization with a single, easy-to-use solution. With Ready PCR Mix, your laboratory can achieve high quality DNA amplification while slashing time and consumption of supplies and hazardous reagents. Ready PCR Mix can:
- Reduce PCR reaction set up and post PCR steps
- Streamline colony screening by eliminating the need for DNA purification
- Eliminate Ethidium Bromide staining and associated waste disposal expenses
The Ready PCR Mix solution includes:
- Extender™ Taq DNA Polymerase
- High fidelity polymerase blend
- Efficient amplification of long and challenging templates (To learn more, click on the features tab of the linked page)
- Reaction Buffer
- MgCl2
- dNTPs
- Density Reagent/Tracking Dye
- Load reaction aliquots directly onto gel
- Non-interfering, fast migrating dye
- Fluorescent DNA Dye
- Based on AMRESCO\’s non-mutagenic and non-toxic EZ-Vision™ formulation
- Immediate visualization of amplified product with no post-run staining or destaining
- Eliminates use of Ethidium Bromide and associated hazardous waste disposal costs
Efficient Amplification of Purified DNA with Ready PCR Mix
Figure 1. Single copy targets were amplified from 50ng of pUC19 plasmid (A) or S. aureus genomic DNA (B) for 35 cycles. 10μl of each PCR reaction was directly loaded onto a 1\% agarose gel and visualized with a Syngene HR gel doc system.
Direct Colony Screening with Ready PCR Mix
Figure 2. 10 distinct colonies were suspended into Ready PCR Mix in separate PCR tubes. 5 μl of the PCR reaction mix was plated on gridded LB plates with Kanamycin for 37ºC overnight growth. After 30 PCR cycles products were screened on a 1\% agarose gel. Colonies 4, 8, 9 and 10 contained the desired sequences. Courtesy of Dr. Irene Lee, Case Western Reserve University, Cleveland, OH
DNA Amplification by Ready PCR Mix Outperforms a Competitor Master Mix
Figure 3. Amplification products from AMRESCO Ready PCR Mix (Lanes 5-8) and a competitor master mix (Lanes 1-4) were resolved on a TAE/1.5\% agarose gel containing Ethidium Bromide. Ethidium Bromide was included in the gel to visualize the products from the competitor master mix which lacks a DNA dye. Lanes 1, 2, 3: Aliquots (5 µl) from triplicate reactions with competitor master mix. Lanes 5,6,7: Aliquots (5 µl) from triplicate reactions with AMRESCO Ready PCR mix. Lanes 4 & 8: Non-template controls (5 µl) from competitor master mix and AMRESCO Ready PCR mix respectively. Data courtesy of Norgen Biotek Corporation.
方形瓶A型
| 产品编号 | 产品名称 | 产品规格 | 产品等级 | 产品价格 |
| 2129 | 方形瓶A型 | 1L | – | – |
| 2128 | 方形瓶A型 | 500mL | – | – |
| 2127 | 方形瓶A型 | 250mL | – | – |
| 2126 | 方形瓶A型 | 100mL | – | – |
| 2125 | 方形瓶A型 | 50mL | – | – |
方形瓶A型

◆优点
有刻度,可以知道溶液量,是由PP制成(盖子是PE制成)
|
容量 |
50mL |
100mL |
250mL |
500mL |
1L |
|
尺寸 |
34×34× 90H mm |
41×41×110H mm |
57×57×126H mm |
71×71×153H mm |
91×91×172H mm |
|
口径 |
17φ mm |
17φ mm |
30φ mm |
43φ mm |
43φ mm |
Q1: 本体、盖子和内盖的材质和耐热温度是什么?
A1: 本体是由聚丙烯制成,耐热温度130℃左右。
盖子和内盖由聚乙烯制成,耐热温度80℃左右。
Q2: 加入1%的硝酸之后,瓶子的颜色变成了粉色,请问是什么原因造成呢?
是不是硝酸导致的?
A2: 苯酚化合物的氧化会引起聚丙烯变色。
● 苯酚化合物的来源
1、树脂中的添加剂(一般等级的PP基本上全部都是由苯酚化合物配制成)
2、包装膜或者纸皮箱的试剂中含有苯酚化合物
● 容易引起变色的情况
1、制品成型后长时间保存
2、在暗处保存
3、在含氮化合物多的空气中保存
4、在与印刷品、包装膜、纸箱等接触的情况下长时间保存
如果变色与苯酚类化合物的氧化有关,促进氧化的硝酸也是原因之一,
如果是与低浓度的硝酸接触一般是不会发生变色,大概是保存的坏境
引起的变色。
Q3: 怎样才能防止变色?
A3: 1、避免接触能够促进苯酚化合物氧化的物质(排气、印刷品)和促进
氧化的坏境(高温高湿)。
2、包装时请用漂白纸或者印刷纸
3、避免进行长期保存
Q4: 请问能够用于金属分析吗?
A4: 不仅这款产品,塑料中都会含有微量的金属,可能会对分析造成影响。
所以请进行实验前先进行空白实验验证。适合金属分析的是聚四氟乙烯
产品,但是圆形的话是由PFA制成,方形的产品是消除了PTFE,使用后
一定要清晰干净。
Phos-tag™ 质谱分析试剂盒
| 产品编号 | 产品名称 | 产品规格 | 产品等级 | 产品价格 |
| 305-93551 | Phos-tag™ Mass Analytical Kit Phos-tag 质谱分析试剂盒 |
1 kit | – | – |
Phos-tag™ Mass Analytical Kit
用于MALDI-TOF/MS检测,提高检测灵敏度!
用于质谱分析的试剂套装。
Phos-tag Mass Analytical Kit 是用于质谱分析的试剂套装,可配套MALDI-TOF/MS使用。可检测磷酸化分子- Phos-tag® 复合物,通常可提高低磷酸化分子的检测灵敏度。
试剂盒内容:
● Phos-tag™ MS-101L 5 mg([C27H29N6O64Zn2]3+ MW:581.4)
● Phos-tag™ MS-101H 5 mg([C27H29N6O68Zn2]3+ MW:589.4)
● Phos-tag™ MS-101N 10 mg([C27H29N6OZn2]3+ MW:584.3)
◆原理:

◆优点、特色:
● CH3COO- 等价结合在Phos-tag™ MS-101上。
● 在溶液中,不含有阴离子的Phos-tag™ MS-101 带有+3价。
● 检测前需制备1mM 的Phos-tag™ MS-101L,MS-101H或者MS-101N( 溶于水)。
◆案例、应用:
【使用例子:检测Phos-tag™ – 磷酸化LPA 复合体】

由于正电荷增大磷酸化LPA 检测灵敏度上升
Phos-tag™ 系列
磷酸化蛋白新方法!
Phos-tag™ 是一种能与磷酸离子特异性结合的功能性分子。它可用于磷酸化蛋白的分离(Phos-tag™ Acrylamide)、Western Blot检测(Phos-tag™ Biotin)、蛋白纯化 (Phos-tag™ Agarose)及质谱分析MALDI-TOF/MS (Phos-tag™ Mass Analytical Kit)。
◆Phos-tag™ 的基本结构:

特点:
与-2价磷酸根离子的亲和性和选择性高于其它阴离子
在pH 5-8的生理环境下生成稳定的复合物
◆原理:
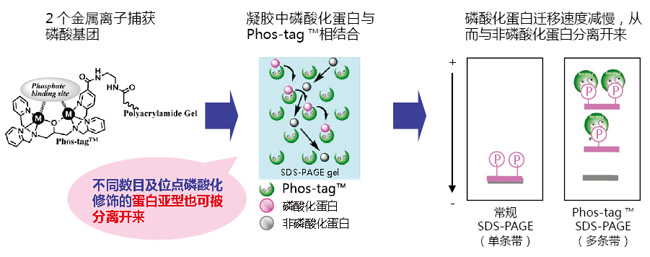
◆相关应用:

◆相关产品:
|
产品名称 |
用 途 |
|
Phos-tag™ Acrylamide |
分离: SDS – PAGE 分离不同磷酸化水平的蛋白 |
|
SuperSep Phos-tag™ |
分离: 预制胶中含有50μM Phos-tag™ Acrylamide |
|
Phos-tag™ Biotin |
检测: 代替 Western Blot 检测中的磷酸化抗体 |
|
Phos-tag™ Agarose |
纯化: 通用柱层析,纯化磷酸化蛋白 |
|
Phos-tag™ Mass Analytical Kit |
分析: 用于质谱 MALDI-TOF/MS 分析,提高磷酸化分子的检测灵敏度 |
phos-tag™由日本广岛大学研究生院医齿药学综合研究科医药分子功能科学研究室开发。
更多产品信息,请点击:http://phos-tag.jp

Phos-tag 第5版说明书

Phos-tag系列 ver 5
Q. Phos-tag™ Mass 用于实验可以使用多少次?
A. 如果每次用量为5μL,至少可以使用1000 次。
Q. 如何选择使用Phos-tag™ MS-101L,Phos-tag™ MS-101H 和Phos-tag™ MS-101N ?
A. Phos-tag™ 101N 含有自然存在的Zn,101L 与101H 分别含有Zn 的同位素64Zn 和68Zn。
请参考以下建议:
摸索条件时使用101N,其中含有多种同位素,结果比较详细;
鉴定磷酸基团是否存在,使用101L 和101H,这些试剂分别包含64Zn 和68Zn。使用这些试剂检测同一个样品
时会产生不同的荷质比。
Q. 如果想测定经过Phos-tag™ SDS-PAGE 分离得到的样品,是否必须要在凝胶消化之前去除Phos-tag™?
A. 没有必要。SDS-PAGE 结束之后根据一般的凝胶消化方法进行操作即可。
Q. 能否用于ESI 质谱?
A. 是的,可以使用。请参考下面的文献,这篇报道使用Phos-tag™ MS-101N 进行ESI-MS 分析。在实验过程
中,使用了中性溶液,若为酸性溶液会导致Phos-tag ™ 分离。
【参考文献】 Anal. Chem. (2008), 80, 2531-2538 (MS-101N ESI-MS)
【参考文献】
· Conversion of graded phosphorylation into switch-like nuclear translocation via autoregulatory mechanisms in ERK signalling[J].Nature communications, 2016, 7,Shindo Y, Iwamoto K, Mouri K, et al.
· PTEN modulates EGFR late endocytic trafficking and degradation by dephosphorylating Rab7[J]. Nature communications, 2016, 7,Shinde S R, Maddika S.
· Feedback control of ErbB2 via ERK-mediated phosphorylation of a conserved threonine in the juxtamembrane domain[J]. Scientific Reports, 2016, 6: 31502,Kawasaki Y, Sakimura A, Park C M, et al.
· Plastid-nucleus communication involves calcium-modulated MAPK signalling[J]. Nature Communications, 2016, 7,Guo H, Feng P, Chi W, et al.
· Sequential domain assembly of ribosomal protein S3 drives 40S subunit maturation[J]. Nature communications, 2016, 7,Mitterer V, Murat G, Réty S, et al.
· Phos-tag analysis of Rab10 phosphorylation by LRRK2: a powerful assay for assessing kinase function and inhibitors[J]. Biochemical Journal, 2016: BCJ20160557,Ito G, Katsemonova K, Tonelli F, et al.
· Analysis of phosphorylation of the myosin targeting subunit of smooth muscle myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. The FASEB Journal, 2016, 30(1 Supplement): 1209.1-1209.1,Walsh M P, MacDonald J A, Sutherland C.
· Using Phos-Tag in Western Blotting Analysis to Evaluate Protein Phosphorylation[J]. Kidney Research: Experimental Protocols, 2016: 267-277,Horinouchi T, Terada K, Higashi T, et al.
· The Abundance of Nonphosphorylated Tau in Mouse and Human Tauopathy Brains Revealed by the Use of Phos-Tag Method[J]. The American journal of pathology, 2016, 186(2): 398-409,Kimura T, Hatsuta H, Masuda-Suzukake M, et al.
· Phos-tag SDS-PAGE resolves agonist-and isoform-specific activation patterns for PKD2 and PKD3 in cardiomyocytes and cardiac fibroblasts[J]. Journal of Molecular and Cellular Cardiology, 2016,Qiu W, Steinberg S F.
· Analysis of phosphorylation of the myosin-targeting subunit of myosin light chain phosphatase by Phos-tag SDS-PAGE[J]. American Journal of Physiology-Cell Physiology, 2016, 310(8): C681-C691,Sutherland C, MacDonald J A, Walsh M P.
· Electrochemical biosensor for protein kinase A activity assay based on gold nanoparticles-carbon nanospheres, phos-tag-biotin and β-galactosidase[J]. Biosensors and Bioelectronics, 2016, 86: 508-515,Zhou Y, Yin H, Li X, et al.
· Validation of Cis and Trans Modes in Multistep Phosphotransfer Signaling of Bacterial Tripartite Sensor Kinases by Using Phos-Tag SDS-PAGE[J]. PloS one, 2016, 11(2): e0148294,Kinoshita-Kikuta E, Kinoshita E, Eguchi Y, et al.
· Phosphopeptide Detection with Biotin-Labeled Phos-tag[J]. Phospho-Proteomics: Methods and Protocols, 2016: 17-29,Kinoshita-Kikuta E, Kinoshita E, Koike T.
· A Phos‐tag SDS‐PAGE method that effectively uses phosphoproteomic data for profiling the phosphorylation dynamics of MEK1[J]. Proteomics, 2016,Kinoshita E, Kinoshita‐Kikuta E, Kubota Y, et al.
· Difference gel electrophoresis of phosphoproteome: U.S. Patent Application 15/004,339[P]. 2016-1-22,Tao W A, Wang L.
· ERK1/2-induced phosphorylation of R-Ras GTPases stimulates their oncogenic potential[J]. Oncogene, 2016,Frémin C, Guégan J P, Plutoni C, et al.
· Microtubules Inhibit E-Cadherin Adhesive Activity by Maintaining Phosphorylated p120-Catenin in a Colon Carcinoma Cell Model[J]. PloS one, 2016, 11(2): e0148574,Maiden S L, Petrova Y I, Gumbiner B M.
· Serine 231 and 257 of Agamous-like 15 are phosphorylated in floral receptacles[J]. Plant Signaling & Behavior, 2016, 11(7): e1199314,Patharkar O R, Macken T A, Walker J C.
· A small molecule pyrazolo [3, 4-d] pyrimidinone inhibitor of zipper-interacting protein kinase suppresses calcium sensitization of vascular smooth muscle[J]. Molecular pharmacology, 2016, 89(1): 105-117,MacDonald J A, Sutherland C, Carlson D A, et al.
· The RNA polymerase II C-terminal domain phosphatase-like protein FIERY2/CPL1 interacts with eIF4AIII and is essential for nonsense-mediated mRNA decay in Arabidopsis[J]. The Plant Cell, 2016: TPC2015-00771-RA,Chen T, Qin T, Ding F, et al.
· Vasorelaxant Effect of 5′-Methylthioadenosine Obtained from Candida utilis Yeast Extract through the Suppression of Intracellular Ca2+ Concentration in Isolated Rat Aorta[J]. Journal of agricultural and food chemistry, 2016, 64(17): 3362-3370,Kumrungsee T, Akiyama S, Saiki T, et al.
· Inhibition of deubiquitinating activity of USP14 decreases tyrosine hydroxylase phosphorylated at Ser19 in PC12D cells[J]. Biochemical and biophysical research communications, 2016, 472(4): 598-602,Nakashima A, Ohnuma S, Kodani Y, et al.
· Actin Tyrosine-53-Phosphorylation in Neuronal Maturation and Synaptic Plasticity[J]. The Journal of Neuroscience, 2016, 36(19): 5299-5313,Bertling E, Englund J, Minkeviciene R, et al.
· AMPK-dependent phosphorylation of lipid droplet protein PLIN2 triggers its degradation by CMA[J]. Autophagy, 2016, 12(2): 432-438,Kaushik S, Cuervo A M.
· Myocardin-related transcription factor a and yes-associated protein exert dual control in G protein-coupled receptor-and RhoA-mediated transcriptional regulation and cell proliferation[J]. Molecular and cellular biology, 2016, 36(1): 39-49,Olivia M Y, Miyamoto S, Brown J H.
· Extensive phosphorylation of AMPA receptors in neurons[J]. Proceedings of the National Academy of Sciences, 2016, 113(33): E4920-E4927,Diering G H, Heo S, Hussain N K, et al.
· The transmembrane region of guard cell SLAC1 channels perceives CO2 signals via an ABA-independent pathway in Arabidopsis[J]. The Plant Cell, 2016, 28(2): 557-567,Yamamoto Y, Negi J, Wang C, et al.
· The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of molecular and cellular cardiology, 2016, 90: 1-10,Kimura T E, Duggirala A, Smith M C, et al.
· Atg13 is essential for autophagy and cardiac development in mice[J]. Molecular and cellular biology, 2016, 36(4): 585-595,Kaizuka T, Mizushima N.
· The ChrSA and HrrSA two-component systems are required for transcriptional regulation of the hemA promoter in Corynebacterium diphtheriae[J]. Journal of Bacteriology, 2016: JB. 00339-16,Burgos J M, Schmitt M P.
· Intergenic Variable-Number Tandem-Repeat Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and Immunity, 2016, 84(7): 2086-2093,Zhu L, Olsen R J, Horstmann N, et al.
· Receptor for advanced glycation end products (RAGE) knockout reduces fetal dysmorphogenesis in murine diabetic pregnancy[J]. Reproductive Toxicology, 2016, 62: 62-70,Ejdesjö A, Brings S, Fleming T, et al.
· Aurora kinase-induced phosphorylation excludes transcription factor RUNX from the chromatin to facilitate proper mitotic progression[J]. Proceedings of the National Academy of Sciences, 2016, 113(23): 6490-6495,Chuang L S H, Khor J M, Lai S K, et al.
· Quantitative phosphoproteomics of protein kinase SnRK1 regulated protein phosphorylation in Arabidopsis under submergence[J]. Journal of experimental botany, 2016: erw107,Cho H Y, Wen T N, Wang Y T, et al.
· Temporal regulation of lipin activity diverged to account for differences in mitotic programs[J]. Current Biology, 2016, 26(2): 237-243,Makarova M, Gu Y, Chen J S, et al.
· Block of CDK1‐dependent polyadenosine elongation of Cyclin B mRNA in metaphase‐i‐arrested starfish oocytes is released by intracellular pH elevation upon spawning[J]. Molecular reproduction and development, 2016, 83(1): 79-87,Ochi H, Aoto S, Tachibana K, et al.
· Mitotic Exit Function of Polo-like Kinase Cdc5 Is Dependent on Sequential Activation by Cdk1[J]. Cell reports, 2016, 15(9): 2050-2062,Rodriguez-Rodriguez J A, Moyano Y, Játiva S, et al.
· PLK2 phosphorylates and inhibits enriched TAp73 in human osteosarcoma cells[J]. Cancer medicine, 2016, 5(1): 74-87,Hu Z B, Liao X H, Xu Z Y, et al.
· Phosphorylated TDP-43 becomes resistant to cleavage by calpain: A regulatory role for phosphorylation in TDP-43 pathology of ALS/FTLD[J]. Neuroscience research, 2016, 107: 63-69,Yamashita T, Teramoto S, Kwak S.
· The Pch2 AAA+ ATPase promotes phosphorylation of the Hop1 meiotic checkpoint adaptor in response to synaptonemal complex defects[J]. Nucleic acids research, 2016: gkw506,Herruzo E, Ontoso D, González-Arranz S, et al.
· An optimized guanidination method for large‐scale proteomic studies[J]. Proteomics, 2016,Ye J, Zhang Y, Huang L, et al.
· Expression and purification of the kinase domain of PINK1 in Pichia pastoris[J]. Protein Expression and Purification, 2016,Wu D, Qu L, Fu Y, et al.
· BRI2 and BRI3 are functionally distinct phosphoproteins[J]. Cellular signalling, 2016, 28(1): 130-144,Martins F, Rebelo S, Santos M, et al.
· Identification of glycoproteins associated with HIV latently infected cells using quantitative glycoproteomics[J]. Proteomics, 2016,Yang W, Jackson B, Zhang H.
· Regulation of Beclin 1 Protein Phosphorylation and Autophagy by Protein Phosphatase 2A (PP2A) and Death-associated Protein Kinase 3 (DAPK3)[J]. Journal of Biological Chemistry, 2016, 291(20): 10858-10866,Fujiwara N, Usui T, Ohama T, et al.
· Regulatory Implications of Structural Changes in Tyr201 of the Oxygen Sensor Protein FixL[J]. Biochemistry, 2016, 55(29): 4027-4035,Yamawaki T, Ishikawa H, Mizuno M, et al.
· Histone demethylase Jmjd3 regulates osteoblast apoptosis through targeting anti-apoptotic protein Bcl-2 and pro-apoptotic protein Bim[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(4): 650-659,Yang D, Okamura H, Teramachi J, et al.
· Analysis of Molecular Species Profiles of Ceramide-1-phosphate and Sphingomyelin Using MALDI-TOF Mass Spectrometry[J]. Lipids, 2016, 51(2): 263-270,Yamashita R, Tabata Y, Iga E, et al.
· Highly sensitive myosin phosphorylation analysis in the renal afferent arteriole[J]. Journal of Smooth Muscle Research, 2016, 52(0): 45-55,Takeya K.
· Functional dissection of the CroRS two-component system required for resistance to cell wall stressors in Enterococcus faecalis[J]. Journal of bacteriology, 2016, 198(8): 1326-1336,Kellogg S L, Kristich C J.
· Regulation of mitogen-activated protein kinase by protein kinase C and mitogen-activated protein kinase phosphatase-1 in vascular smooth muscle[J]. American Journal of Physiology-Cell Physiology, 2016, 310(11): C921-C930,Trappanese D M, Sivilich S, Ets H K, et al.
· ModProt: a database for integrating laboratory and literature data about protein post-translational modifications[J]. Journal of Electrophoresis, 2016, 60(1): 1-4,Kimura Y, Toda T, Hirano H.
· The C-ETS2-TFEB Axis Promotes Neuron Survival under Oxidative Stress by Regulating Lysosome Activity[J]. Oxidative medicine and cellular longevity, 2016,Ma S, Fang Z, Luo W, et al.
· Essential role of the PSI–LHCII supercomplex in photosystem acclimation to light and/or heat conditions by state transitions[J]. Photosynthesis Research, 2016: 1-10,Marutani Y, Yamauchi Y, Higashiyama M, et al.
· Identification of a redox-modulatory interaction between selenoprotein W and 14-3-3 protein[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2016, 1863(1): 10-18,Jeon Y H, Ko K Y, Lee J H, et al.
· Effects of hydrogen sulfide on the heme coordination structure and catalytic activity of the globin-coupled oxygen sensor AfGcHK[J]. BioMetals, 2016, 29(4): 715-729,Fojtikova V, Bartosova M, Man P, et al.
· Identification and functional analysis of phosphorylation in Newcastle disease virus phosphoprotein[J]. Archives of virology, 2016: 1-14,Qiu X, Zhan Y, Meng C, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports, 2016, 6: 54-62,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· Profiling DNA damage-induced phosphorylation in budding yeast reveals diverse signaling networks[J]. Proceedings of the National Academy of Sciences, 2016: 201602827,Zhou C, Elia A E H, Naylor M L, et al.
· Unexpected properties of sRNA promoters allow feedback control via regulation of a two-component system[J]. Nucleic Acids Research, 2016: gkw642,Brosse A, Korobeinikova A, Gottesman S, et al.
· Evolution of ZnII–Macrocyclic Polyamines to Biological Probes and Supramolecular Assembly[J]. Macrocyclic and Supramolecular Chemistry: How Izatt-Christensen Award Winners Shaped the Field, 2016: 415,Kimura E, Koike T, Aoki S.
· Phosphopeptide Enrichment Using Various Magnetic Nanocomposites: An Overview[J]. Phospho-Proteomics: Methods and Protocols, 2016: 193-209,Batalha Í L, Roque A C A.
· In vivo phosphorylation of a peptide tag for protein purification[J]. Biotechnology letters, 2016, 38(5): 767-772,Goux M, Fateh A, Defontaine A, et al.
· Regulation of cell reversal frequency in Myxococcus xanthus requires the balanced activity of CheY‐like domains in FrzE and FrzZ[J]. Molecular microbiology, 2016,Kaimer C, Zusman D R.
· Elevation of cortical serotonin transporter activity upon peripheral immune challenge is regulated independently of p38 mitogen‐activated protein kinase activation and transporter phosphorylation[J]. Journal of neurochemistry, 2016, 137(3): 423-435,Schwamborn R, Brown E, Haase J.
· The Yeast Cyclin-Dependent Kinase Routes Carbon Fluxes to Fuel Cell Cycle Progression[J]. Molecular cell, 2016, 62(4): 532-545,Ewald J C, Kuehne A, Zamboni N, et al.
· Two Degradation Pathways of the p35 Cdk5 (Cyclin-dependent Kinase) Activation Subunit, Dependent and Independent of Ubiquitination[J]. Journal of Biological Chemistry, 2016, 291(9): 4649-4657,Takasugi T, Minegishi S, Asada A, et al.
· Increased level of phosphorylated desmin and its degradation products in heart failure[J]. Biochemistry and Biophysics Reports. 2016,Bouvet M, Dubois-Deruy E, Alayi T D, et al.
· a high‐affinity LCO‐binding protein of Medicago truncatula, interacts with LYK3, a key symbiotic receptor[J]. FEBS letters, 2016, 590(10): 1477-1487,Fliegmann J, Jauneau A, Pichereaux C, et al. LYR3,
· Nek1 Regulates Rad54 to Orchestrate Homologous Recombination and Replication Fork Stability[J]. Molecular Cell, 2016,Spies J, Waizenegger A, Barton O, et al.
· PhostagTM-gel retardation and in situ thylakoid kinase assay for determination of chloroplast protein phosphorylation targets[J]. Endocytobiosis and Cell Research, 2016, 27(2): 62-70,Dytyuk Y, Flügge F, Czarnecki O, et al.
· Luteinizing Hormone Causes Phosphorylation and Activation of the cGMP Phosphodiesterase PDE5 in Rat Ovarian Follicles, Contributing, Together with PDE1 Activity, to the Resumption of Meiosis[J]. Biology of reproduction, 2016: biolreprod. 115.135897,Egbert J R, Uliasz T F, Shuhaibar L C, et al.
· Newby, AC, & Bond, M.(2016). The Hippo pathway mediates inhibition of vascular smooth muscle cell proliferation by cAMP[J]. Journal of Molecular and Cellular Cardiology, 2016, 90: 1-10,Kimura-Wozniak T, Duggirala A, Smith M C, et al. G.
· Yeast lacking the amphiphysin family protein Rvs167 is sensitive to disruptions in sphingolipid levels[J]. The FEBS Journal, 2016, 283(15): 2911-2928,Toume M, Tani M.
· Regulation of CsrB/C sRNA decay by EIIAGlc of the phosphoenolpyruvate: carbohydrate phosphotransferase system[J]. Molecular microbiology, 2016, 99(4): 627-639,Leng Y, Vakulskas C A, Zere T R, et al.
· The Late S-Phase Transcription Factor Hcm1 Is Regulated through Phosphorylation by the Cell Wall Integrity Checkpoint[J]. Molecular and cellular biology, 2016: MCB. 00952-15,Negishi T, Veis J, Hollenstein D, et al.
· Validation of chemical compound library screening for transcriptional co‐activator with PDZ‐binding motif inhibitors using GFP‐fused transcriptional co‐activator with PDZ‐binding motif[J]. Cancer science, 2016, 107(6): 791-802,Nagashima S, Maruyama J, Kawano S, et al.
· ULK1/2 Constitute a Bifurcate Node Controlling Glucose Metabolic Fluxes in Addition to Autophagy[J]. Molecular cell, 2016, 62(3): 359-370,Li T Y, Sun Y, Liang Y, et al.
· Spatiotemporal dynamics of Oct4 protein localization during preimplantation development in mice[J]. Reproduction, 2016: REP-16-0277,Fukuda A, Mitani A, Miyashita T, et al.
· The tandemly repeated NTPase (NTPDase) from Neospora caninum is a canonical dense granule protein whose RNA expression, protein secretion and phosphorylation coincides with the tachyzoite egress[J]. Parasites & Vectors, 2016, 9(1): 1,Pastor-Fernández I, Regidor-Cerrillo J, Álvarez-García G, et al.
· Interaction Analysis of a Two-Component System Using Nanodiscs[J]. PloS one, 2016, 11(2): e0149187,Hörnschemeyer P, Liss V, Heermann R, et al.
· Constitutive Activation of PINK1 Protein Leads to Proteasome-mediated and Non-apoptotic Cell Death Independently of Mitochondrial Autophagy[J]. Journal of Biological Chemistry, 2016, 291(31): 16162-16174,Akabane S, Matsuzaki K, Yamashita S, et al.
· p38β Mitogen-Activated Protein Kinase Modulates Its Own Basal Activity by Autophosphorylation of the Activating Residue Thr180 and the Inhibitory Residues Thr241 and Ser261[J]. Molecular and cellular biology, 2016, 36(10): 1540-1554,Beenstock J, Melamed D, Mooshayef N, et al.
· Lysophosphatidylcholine acyltransferase 1 protects against cytotoxicity induced by polyunsaturated fatty acids[J]. The FASEB Journal, 2016, 30(5): 2027-2039,Akagi S, Kono N, Ariyama H, et al.
· Characterization of a herpes simplex virus 1 (HSV-1) chimera in which the Us3 protein kinase gene is replaced with the HSV-2 Us3 gene[J]. Journal of virology, 2016, 90(1): 457-473,Shindo K, Kato A, Koyanagi N, et al.
· Generation of phospho‐ubiquitin variants by orthogonal translation reveals codon skipping[J]. FEBS letters, 2016, 590(10): 1530-1542,George S, Aguirre J D, Spratt D E, et al.
· Evolution of KaiC-Dependent Timekeepers: A Proto-circadian Timing Mechanism Confers Adaptive Fitness in the Purple Bacterium Rhodopseudomonas palustris[J]. PLoS Genet, 2016, 12(3): e1005922,Ma P, Mori T, Zhao C, et al.
· Phosphorylation of Bni4 by MAP kinases contributes to septum assembly during yeast cytokinesis[J]. FEMS Yeast Research, 2016, 16(6): fow060,Pérez J, Arcones I, Gómez A, et al.
· Alteration of Antiviral Signalling by Single Nucleotide Polymorphisms (SNPs) of Mitochondrial Antiviral Signalling Protein (MAVS)[J]. PloS one, 2016, 11(3): e0151173,Xing F, Matsumiya T, Hayakari R, et al.
· Arm-in-arm response regulator dimers promote intermolecular signal transduction[J]. Journal of bacteriology, 2016, 198(8): 1218-1229,Baker A W, Satyshur K A, Morales N M, et al.
· The lsh/ddm1 homolog mus-30 is required for genome stability, but not for dna methylation in neurospora crassa[J]. PLoS Genet, 2016, 12(1): e1005790,Basenko E Y, Kamei M, Ji L, et al.
· Fine tuning chloroplast movements through physical interactions between phototropins[J]. Journal of Experimental Botany, 2016: erw265,Sztatelman O, Łabuz J, Hermanowicz P, et al.
· Characterization of the Neospora caninum NcROP40 and NcROP2Fam-1 rhoptry proteins during the tachyzoite lytic cycle[J]. Parasitology, 2016, 143(01): 97-113,Pastor-Fernandez I, Regidor-Cerrillo J, Jimenez-Ruiz E, et al.
· Transcriptional Profile during Deoxycholate-Induced Sporulation in a Clostridium perfringens Isolate Causing Foodborne Illness[J]. Applied and environmental microbiology, 2016, 82(10): 2929-2942,Yasugi M, Okuzaki D, Kuwana R, et al.
· Timely Closure of the Prospore Membrane Requires SPS1 and SPO77 in Saccharomyces cerevisiae[J]. Genetics, 2016: genetics. 115.183939,Paulissen S M, Slubowski C J, Roesner J M, et al.
· DDK dependent regulation of TOP2A at centromeres revealed by a chemical genetics approach[J]. Nucleic Acids Research, 2016: gkw626,Wu K Z L, Wang G N, Fitzgerald J, et al.
· OVATE Family Protein 8 Positively Mediates Brassinosteroid Signaling through Interacting with the GSK3-like Kinase in Rice[J]. PLoS Genet, 2016, 12(6): e1006118,Yang C, Shen W, He Y, et al.
· Epithelial Sel1L is required for the maintenance of intestinal homeostasis[J]. Molecular biology of the cell, 2016, 27(3): 483-490, Sun S, Lourie R, Cohen S B, et al.
· Effect of Sodium Dodecyl Sulfate Concentration on Supramolecular Gel Electrophoresis[J]. ChemNanoMat, 2016,Tazawa S, Kobayashi K, Yamanaka M.
· Intergenic VNTR Polymorphism Upstream of rocA Alters Toxin Production and Enhances Virulence in Streptococcus pyogenes[J]. Infection and immunity, 2016: IAI. 00258-16,Zhu L, Olsen R J, Horstmann N, et al.
· Ajuba Phosphorylation by CDK1 Promotes Cell Proliferation and Tumorigenesis[J]. Journal of Biological Chemistry, 2016: jbc. M116. 722751,Chen X, Stauffer S, Chen Y, et al.
· Editorial: International Plant Proteomics Organization (INPPO) World Congress 2014[J]. Frontiers in Plant Science, 2016, 7,Heazlewood J L, Jorrín-Novo J V, Agrawal G K, et al.
· Phosphoinositide kinase signaling controls ER-PM cross-talk[J]. Molecular biology of the cell, 2016, 27(7): 1170-1180,Omnus D J, Manford A G, Bader J M, et al.
· A multiple covalent crosslinked soft hydrogel for bioseparation[J]. Chemical Communications, 2016, 52(15): 3247-3250,Liu Z, Fan L, Xiao H, et al.
· Advances in crop proteomics: PTMs of proteins under abiotic stress[J]. Proteomics, 2016, 16(5): 847-865,Wu X, Gong F, Cao D, et al.
· Cyclin-Dependent Kinase Co-Ordinates Carbohydrate Metabolism and Cell Cycle in S. cerevisiae[J]. Molecular cell, 2016, 62(4): 546-557,Zhao G, Chen Y, Carey L, et al.
· Carbon Monoxide Gas Is Not Inert, but Global, in Its Consequences for Bacterial Gene Expression, Iron Acquisition, and Antibiotic Resistance[J]. Antioxidants & redox signaling, 2016,Wareham L K, Begg R, Jesse H E, et al.
· Two-layer regulation of PAQR3 on ATG14-linked class III PtdIns3K activation upon glucose starvation[J]. Autophagy, 2016: 1-2,Xu D, Wang Z, Chen Y.
· Regulation of sphingolipid biosynthesis by the morphogenesis checkpoint kinase Swe1[J]. Journal of Biological Chemistry, 2016, 291(5): 2524-2534,Chauhan N, Han G, Somashekarappa N, et al.
· PAX5 tyrosine phosphorylation by SYK co-operatively functions with its serine phosphorylation to cancel the PAX5-dependent repression of BLIMP1: A mechanism for antigen-triggered plasma cell differentiation[J]. Biochemical and biophysical research communications, 2016, 475(2): 176-181,Inagaki Y, Hayakawa F, Hirano D, et al.
· A Combined Computational and Genetic Approach Uncovers Network Interactions of the Cyanobacterial Circadian Clock[J]. Journal of Bacteriology, 2016: JB. 00235-16,Boyd J S, Cheng R R, Paddock M L, et al.
· HuR mediates motility of human bone marrow-derived mesenchymal stem cells triggered by sphingosine 1-phosphate in liver fibrosis[J]. Journal of Molecular Medicine, 2016: 1-14,Chang N, Ge J, Xiu L, et al.
· Combined replacement effects of human modified β-hexosaminidase B and GM2 activator protein on GM2 gangliosidoses fibroblasts[J]. Biochemistry and Biophysics Reports, 2016,Kitakaze K, Tasaki C, Tajima Y, et al.
· Roseotoxin B Improves Allergic Contact Dermatitis through a Unique Anti-inflammatory Mechanism Involving Excessive Activation of Autophagy in Activated T-Lymphocytes[J]. Journal of Investigative Dermatology, 2016,Wang X, Hu C, Wu X, et al.
References on Phos-tag™ Chemistry
-
Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry of phosphorylated compounds using a novel phosphate capture molecule, Rapid Communications of Mass Spectrometry, 17, 2075-2081 (2003), H. Takeda, A. Kawasaki, M. Takahashi, A. Yamada, and T. Koike
-
Recognition of phosphate monoester dianion by an alkoxide-bridged dinuclear zinc (II) complex, Dalton Transactions, 1189-1193 (2004), E. Kinoshita, M. Takahashi, H. Takeda, M. Shiro, and T. Koike
-
Quantitative analysis of lysophosphatidic acid by time-of-flight mass spectrometry using a phosphate capture molecule, Journal of Lipid Research, 45, 2145-2150 (2004), T. Tanaka, H. Tsutsui, K. Hirano, T. Koike, A. Tokumura, and K. Satouchi
-
Production of 1,2-Didocosahexaenoyl Phosphatidylcholine by Bonito Muscle Lysophosphatidylcholine/Transacylase, Journal of Biochemistry,136, 477-483 (2004), K. Hirano, H. Matsui, T. Tanaka, F. Matsuura, K. Satouchi, and T. Koike
-
Novel immobilized zinc(II) affinity chromatography for phosphopeptides and phosphorylated proteins, Journal of Separation Science, 28, 155-162 (2005), E. Kinoshita, A. Yamada, H. Takeda, E. Kinoshita-Kikuta, and T. Koike
-
Detection and Quantification of On-Chip Phosphorylated Peptides by Surface Plasmon Resonance Imaging Techniques Using a Phosphate Capture Molecule, Analytical Chemistry, 77, 3979-3985 (2005), K. Inamori, M. Kyo, Y. Nishiya, Y. Inoue, T. Sonoda, E. Kinoshita, T. Koike, and Y. Katayama
-
Phosphate-binding tag: A new tool to visualize phosphorylated proteins, Molecular & Cellular Proteomics, 5, 749-757 (2006), E. Kinoshita, E. Kinoshita-Kikuta, K. Takiyama, and T. Koike
-
Enrichment of phosphorylated proteins from cell lysate using phosphate-affinity chromatography at physiological pH, Proteomics, 6, 5088-5095 (2006), E. Kinoshita-Kikuta, E. Kinoshita, A. Yamada, M. Endo, and T. Koike
-
Separation of a phosphorylated histidine protein using phosphate affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 360, 160-162 (2007), S. Yamada, H. Nakamura, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and Y. Shiro
-
Label-free kinase profiling using phosphate-affinity polyacrylamide gel electrophresis, Molecular & Cellular Proteomics, 6, 356-366 (2007), E. Kinoshita-Kikuta, Y. Aoki, E. Kinoshita, and T. Koike
-
A SNP genotyping method using phosphate-affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 361, 294-298 (2007), E. Kinoshita, E. Kinoshita-Kikuta, and T. Koike (The phosphate group at DNA-terminal is efficiently captured by Zn2+.Phos-tag.)
-
Identification on Membrane and Characterization of Phosphoproteins Using an Alkoxide-Bridged Dinuclear Metal Complex as a Phosphate-Binding Tag Molecule, Journal of Biomolecular Techniques, 18, 278-286 (2007), T. Nakanishi, E. Ando, M. Furuta, E. Kinoshita, E. Kikuta-Kinoshita, T. Koike, S. Tsunasawa, and O. Nishimura
-
A mobility shift detection method for DNA methylation analysis using phosphate affinity polyacrylamide gel electrophoresis, Analytical Biochemistry, 378, 102-104 (2008), E. Kinoshita-Kikuta, E. Kinoshita, and T. Koike
-
Separation of phosphoprotein isotypes having the same number of phosphate groups using phosphate- affinity SDS-PAGE, Proteomics, 8, 2994-3003 (2008), E. Kinoshita, E. Kinoshita-Kikuta, M. Matsubara, S. Yamada, H. Nakamura, Y. Shiro, Y. Aoki, K. Okita, and T. Koike
-
FANCI phosphorylation functions as a molecular switch to turn on the Fanconi anemia pathway, Nature Structural & Molecular Biology, 15, 1138-1146 (2008), M. Ishiai, H. Kitao, A. Smogorzewska, J. Tomida, A. Kinomura, E. Uchida, A. Saberi, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, S. Tashiro, S. J. Elledge, and M. Takata
-
Two-dimensional phosphate affinity gel electrophoresis for the analysis of phosphoprotein isotypes , Electrophoresis, 30, 550-559 (2009), E. Kinoshita, E. Kinoshita-Kikuta, M. Matsubara, Y. Aoki, S. Ohie, Y. Mouri, and T. Koike
-
Formation of lysophosphatidic acid, a wound-healing lipid, during digestion of cabbage leaves, Bioscience, Biotechnology, and Biochemistry,73, 1293-300 (2009), T. Tanaka, G. Horiuchi, M. Matsuoka, K. Hirano, A. Tokumura, T. Koike, and K. Satouchi
-
A Phos-tag-based fluorescence resonance energy transfer system for the analysis of the dephosphorylation of phosphopeptides, Analytical Biochemistry, 388, 235-241, (2009), K. Takiyama, E. Kinoshita, E. Kinoshita-Kikuta, Y. Fujioka, Y. Kubo, and T. Koike
-
Phos-tag beads as an immunoblotting enhancer for selective detection of phosphoproteins in cell lysates, Analytical Biochemistry, 389, 83-85, (2009), E. Kinoshita-Kikuta, E. Kinoshita, and T. Koike
-
Mobility shift detection of phosphorylation on large proteins using a Phos-tag SDS-PAGE gel strengthened with agarose, Proteomics, 9, 4098- 4101 (2009), E. Kinoshita, E. Kinoshita-Kikuta, H. Ujihara, and T. Koike
-
Separation and detection of large phosphoproteins using Phos-tag SDS-PAGE, Nature Protocols, 4, 1513-1521 (2009), E. Kinoshita, E. Kinoshita-Kikuta, and T. Koike
-
A clean-up technology for the simultaneous determination of lysophosphatidic acid and sphingosine-1-phosphate by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry using a phosphate-capture molecule, Phos-tag, Rapid Communications in Mass Spectrometry, 24, 1075-1084 (2010), J. Morishige, M. Urikura, H. Takagi, K. Hirano, T. Koike, T. Tanaka, and K. Satouchi
-
Genotyping and mapping assay of single-nucleotide polymorphisms in CYP3A5 using DNA-binding zinc(II) complexes, Clinical Biochemistry, 43, 302-306 (2010), E. Kinoshita, E. Kinoshita-Kikuta, H. Nakashima, and T. Koike
-
The DNA-binding activity of mouse DNA methyltransferase 1 is ragulated phosphorylation with casein kinase 1σ/ε, Biochemical Journal, 427, 489-497 (2010), Y. Sugiyama, N. Hatano, N. Sueyoshi, I. Suetake, S. Tajima, E. Kinoshita, E. Kinoshita-Kikuta, T. Koike, and I. Kameshita
to Page top